Страница: 9/13
Дополнительным механизмом, ответственным за вывод ионов кальция из цитоплазмы, является натрий-кальциевый обменник, который выводит Са2+, используя энергию натриевого электрохимического градиента. Наличие Na+- Са2+ обменника было показано в различных типах возбудимых и невозбудимых клеток; в клетках нервной системы он был обнаружен в конце 60-х годов (9). В нейронах моллюска, помещенных в среду с пониженным натрием (т.е. с обратным натриевым градиентом), наблюдалось увеличение [Ca2+]i, что является результатом работы обменника в инвертированной форме. Однако, вклад Na+- Са2+ обменника в регуляцию [Ca2+]i в нейронах млекопитающих до сих пор не оценен. В некоторых работах было показано, что обменник принимает незначительное участие в удалении цитоплазматического Са2+, в то время как в других работах представлены данные о том, что обменник играет существенную роль в переносе Са2+ через мембрану (57).
Кроме быстрого связывания цитозольного Са2+ внутриклеточными Са2+-связывающими белками, ионы кальция, попадающие в цитозоль, могут аккумулироваться аппаратом Гольджи или клеточным ядром, захватываться митохондриальными Са2+ депо, имеющими достаточно невысокое сродство к Са2+, или быстрыми депо, связанными с ЭР или СР, имеющими высокое сродство к Са2+. Однако если [Ca2+]i превышает 0,5 мкмоль/л, наблюдается существенное перераспределение [Ca2+]i в область митохондрий. Буферные системы митохондрий принимают участие в удалении избыточного Са2+ из цитоплазмы в клетках кишечника, некоторых типах нервных клеток (59) и в секреторных клетках после повышения [Ca2+]i, стимулированного агонистами. Связывание кальция митохондриями обеспечивается активностью систем, расположенных на внутренней митохондриальной мембране. Са2+ поступает в митохондрии по электрохимическому градиенту; разность потенциалов, обеспечивающая транспорт кальция, создается переносом электронов во время клеточного дыхания и связанного с ним переносом протонов. Перенос электронов по дыхательной цепи является основным механизмом, обеспечивающим энергетику транспорта кальция. Подавление дыхательной цепи карбонил-цианид-м-хлорофенил-гидразоном (СССР) эффективно блокирует аккумуляцию кальция митохондриями (41).
Последние исследования показали, что АТФ занимает прочное место в ряду нейромедиаторов центральной и периферической нервной систем (Burnstock 1990). Не вызывает сомнения, что АТФ является не только важнейшим внутриклеточным метаболитом, но и служит важным объектом межклеточного взаимодействия.
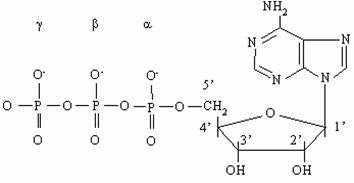 |
АТФ (см. рис.1) представляет собой нуклеотид и как всякий нуклеотид состоит из трех компонентов: азотистого основания, сахара пентозы и фосфата. В качестве азотистого основания в нуклеотидах присутствуют производные пурина и пиримидина. Фосфаты соединены в полифосфатную цепь, количество которых в естественных нуклеотидах не превышает трех. Однако синтезированы нуклеотиды, содержащие линейные цепи из более чем 3-х фосфатов, к примеру аденозинтетра- и аденозинпентафосфаты.
Названия нуклеотидов, содержащих в качестве сахара рибозу, складываются из названия соответствующего нуклеозида, приставки, обозначающей количество фосфатных групп в нуклеотиде и слова фосфат. Для наиболее распространенных нуклеотидов приняты сокращенные названия, например АТФ для аденозинтрифосфата, ГТФ - для гуанозинтрифосфата, ИМФ - инозинмонофосфата.
Реферат опубликован: 12/11/2009