Страница: 4/13
41. Nicholls D.G. (1985) A role for the mitochondria in the protection of the cell against calcium overload. Prog. Brain Res. 63, 97‑106.
42. Pintor, J., Diaz-Rey, M. A., Torres, M., Miras-Portugal, M. T. (1992) Presence of diadenosine polyphosphates-Ap4A and Ap5A-in rat brain synaptic terminals. Ca2+-dependent release evoked by 4-aminopyridine and veratridine. Neurosci. Lett. 136: 141-144
43. Ribeiro, J. A., Sebastiao, A. M. (1986) Adenosine receptors and calcium: basis for proposing a third (A3) adenosine receptor. Prog. Neyrobiol. 26: 179-209
44. Rios E. and Pizarro C. (1991) Voltage‑sensor of excitation‑contraction coupling in skeletal muscle. Physiol. Rev. 76, 849 ‑ 908
45. Ross C. A., Danoff S. K., Schell M. J., Snyder S. H. and Ullrich A. (1992). Three additional inositol 1,4,5‑trisphosphate receptors: Molecular cloning and differential localization in brain and peripheral tissues. Proc. Natl. Acad. Sci. U.S.A. 89, 4265‑4269.
46. S.Kirischuk, N.Voitenko, P.Kostyuk, A.Verkhratsky (1995) Calcium signalling in granule neurones studied in cerebellar slices // Cell Calcium v.18, 464-476
47. S.Kirischuk, N.Voitenko, P.Kostyuk, A.Verkhratsky (1996) Age-associated Changes of Citoplasmic Calcium Homeostasis in Cerebellar Granule Neurones in situ: Investigation on Thin Cerebellar Slices. // Experimental Gerontology
|
|
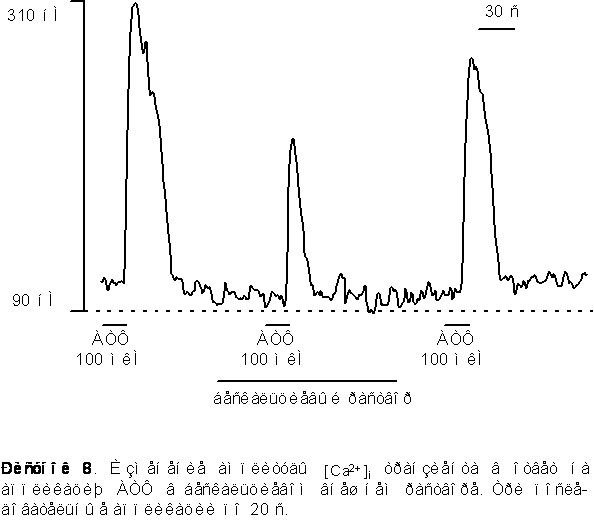
1. Для исследования способности внутриклеточного Ca2+ депо к перезаполнению мы последовательно апплицировали АТФ несколько раз с интервалом в 60 секунд в обычном растворе и в растворе не содержащем Ca2+ . Как видно из рисунка 9 вторая аппликация АТФ вызывала Ca2+ транзиент меньшей (на 30% ± 4%) амплитуды по сравнению с первой (контрольной) аппликацией. Последующие аппликации АТФ разделенные 60 ти секундным интервалом не вызывали заметного уменьшения амплитуды [Ca2+]i транзинета по сравнению со второй аппликацией в стандартном растворе
|
|

В бескальциевом растворе - наоборот, вторая аппликация АТФ вызывала значительное уменьшение Ca2+ транзиента на 40% ± 5% от контроля. Последующие аппликации АТФ в бескальциевом растворе приводили к последовательному уменьшению амплитуды [Ca2+]i транзиента и после двух - трех последовательных аппликаций сигнал исчезал вообще (рисунок 10). Такое уменьшение вероятно обусловлено истощением внутриклеточных депо.
|
|

1. Ингибирование захвата Ca2+ эндоплазматическим ретикулом тапсигаргином (спецефическим блокатором Ca2+ насоса эндоплазматического ретикулума) уменьшало [Ca2+]i транзиенты, вызванные приложением АТФ, на 63% ± 5% для концентрации АТФ 100 мкМ АТФ (рис. 11). Как видно из рисунка 11 приложение тапсигаргина не влияло на базальный уровень Ca2+ в клетке.
1. Приложение кадмия в концентрации 50 мкМ, верапамила в концентрации 100 мкМ и 50 мкМ никеля обратимо уменьшало амплитуду [Ca2+]i транзиента вызванного приложением 100 мкМ АТФ на 35%, 20%, и 15% сответственно. Данные представлены на рисунке 12.
|
|

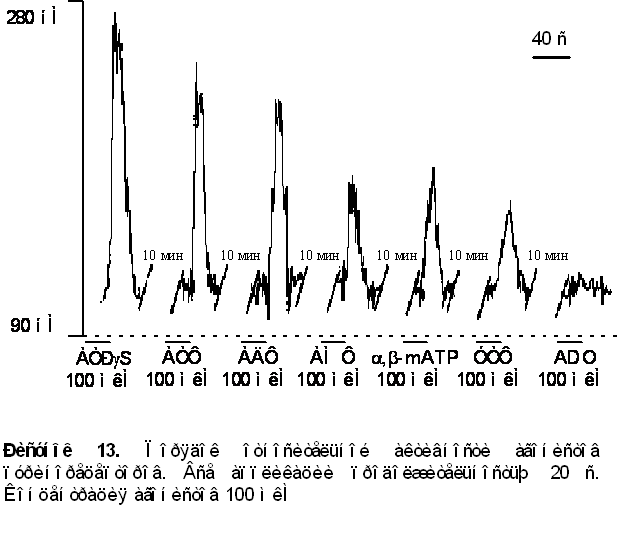
1. Приложение различных агонистов пуринорецепторов вызывало различные по амплитуде ответы. Порядок относительной активности лигандов для данного объекта был следующим ATPgS > ATФ = AДФ >> a,b-methylene ATP » AMФ > УТФ>> аденозин (ADO), данные преставлены на рисунке 13.
|
|
Реферат опубликован: 12/11/2009